Фотосинтез. Общее уравнение фотосинтеза
Химическое уравнение процесса фотосинтеза в общем можно представить в следующем виде:
6СО 2 + 6Н 2 О + Qсвета → С 6 Н 12 О 6 + 6О 2 .
Фотосинтез – процесс, при котором происходит поглощение электромагнитной энергии солнца хлорофиллом и вспомогательными пигментами и превращение её в химическую энергию, поглощение углекислого газа из атмосферы, восстановление его в органические соединения и возвращение кислорода в атмосферу.
В процессе фотосинтеза из простых неорганических соединений (СО 2 , Н 2 О) строятся различные органические соединения. В результате происходит перестройка химических связей: вместо связей С – О и Н – О возникают связи C – C и C – H, в которых электроны занимают более высокий энергетический уровень. Таким образом, богатые энергией органические вещества, которыми питаются и за счет которых получают энергию (в процессе дыхания) животные и человек, первоначально создаются в зеленом листе. Можно сказать, что практически вся живая материя на Земле является результатом фотосинтетической деятельности.
Датой открытия процесса фотосинтеза можно считать 1771 г. Английский ученый Дж. Пристли обратил внимание на изменение состава воздуха вследствие жизнедеятельности животных. В присутствии зеленых растений воздух вновь становился пригодным как для дыхания, так и для горения. В дальнейшем работами ряда ученых (Я. Ингенгауз, Ж. Сенебье, Т. Соссюр, Ж.Б. Буссенго) было установлено, что зеленые растения из воздуха поглощают С0 2 , из которого при участии воды на свету образуется органическое вещество. Именно этот процесс в 1877 г. немецкий ученый В. Пфеффер назвал фотосинтезом. Большое значение для раскрытия сущности фотосинтеза имел закон сохранения энергии, сформулированный Р. Майером. В 1845 г. Р. Майер выдвинул предположение, что энергия, используемая растениями, - это энергия Солнца, которую растения в процессе фотосинтеза превращают в химическую энергию. Это положение было развито и экспериментально подтверждено в исследованиях замечательного русского ученого К.А. Тимирязева.
Фотосинтез включает как световые, так и темновые реакции. Был проведен ряд экспериментов, доказывающих, что в процессе фотосинтеза происходят не только реакции, идущие с использованием энергии света, но и темновые, не требующие непосредственного участия энергии света. Можно привести следующие доказательства существования темновых реакций в процессе фотосинтеза:
1) фотосинтез ускоряется с повышением температуры. Отсюда прямо следует, что какие-то этапы этого процесса непосредственно не связаны с использованием энергии света. Особенно резко зависимость фотосинтеза от температуры проявляется при высоких интенсивностях света. По-видимому, в этом случае скорость фотосинтеза лимитируется именно темновыми реакциями;
2) эффективность использования энергии света в процессе фотосинтеза оказалась выше при прерывистом освещении. При этом для более эффективного использования энергии света длительность темновых промежутков должна значительно превышать длительность световых.
Пигменты фотосинтеза
Для того чтобы свет мог оказывать влияние на растительный организм и, в частности, быть использованным в процессе фотосинтеза, необходимо его поглощение фоторецепторами-пигментами. Пигменты - это окрашенные вещества. Пигменты поглощают свет определенной длины волны. Непоглощенные участки солнечного спектра отражаются, что и обусловливает окраску пигментов. Так, зеленый пигмент хлорофилл поглощает красные и синие лучи, тогда как зеленые лучи в основном отражаются. Видимая часть солнечного спектра включает длины волн от 400 до 700 нм. Вещества, поглощающие весь видимый участок спектра, кажутся черными.
Пигменты, сконцентрированные в пластидах, можно разделить на три группы: хлорофиллы, каротиноиды, фикобилины .
К группе хлорофиллов относят органические соединения, которые содержат 4 пиррольных кольца, соединённых атомами магния и имеющие зелёную окраску.
В настоящее время известно около десяти хлорофиллов. Они отличаются по химическому строению, окраске, распространению среди живых организмов. У всех высших растений содержатся хлорофиллы а и b. Хлорофилл с обнаружен в диатомовых водорослях, хлорофилл d - в красных водорослях.
Основными пигментами, без которых фотосинтез не идет, являются хлорофилл а для зеленых растений и бактериохлорофиллы для бактерий. Впервые точное представление о пигментах зеленого листа высших растений было получено благодаря работам крупнейшего русского ботаника М.С. Цвета (1872-1919). Он разработал новый хроматографический метод разделения веществ и выделил пигменты листа в чистом виде.
Хроматографический метод разделения веществ основан на их различной способности к адсорбции. Метод этот получил широкое применение. М.С. Цвет пропускал вытяжку из листа через стеклянную трубку, заполненную порошком - мелом или сахарозой (хроматографическую колонку). Отдельные компоненты смеси пигментов различались по степени адсорбируемости и передвигались с разной скоростью, в результате чего они концентрировались в разных зонах колонки. Разделяя колонку на отдельные части (зоны) и используя соответствующую систему растворителей, можно было выделить каждый пигмент. Оказалось, что листья высших растений содержат хлорофилл а и хлорофилл b, а также каротиноиды (каротин, ксантофилл и др.). Хлорофиллы, так же как и каротиноиды, нерастворимы в воде, но хорошо растворимы в органических растворителях. Хлорофиллы а и b различаются по цвету: хлорофилл а имеет сине-зеленый оттенок, а хлорофилл b - желто-зеленый. Содержание хлорофилла а в листе примерно в три раза больше по сравнению с хлорофиллом b.
Каротиноиды - это желтые и оранжевые пигменты алифатического строения, производные изопрена. Каротиноиды содержатся во всех высших растениях и у многих микроорганизмов. Это самые распространенные пигменты с разнообразными функциями. Каротинойды, содержащие кислород, получили название ксантофиллы. Основными представителями каротиноидов у высших растений являются два пигмента - каротин (оранжевый) и ксантофилл (желтый). В отличие от хлорофиллов каротиноиды не поглощают красные лучи, а также не обладают способностью к флуоресценции. Подобно хлорофиллу каротиноиды в хлоропластах и хроматофорах находятся в виде нерастворимых в воде комплексов с белками. Каротиноиды, поглощая определенные участки солнечного спектра, передают энергию этих лучей на молекулы хлорофилла. Тем самым они способствуют использованию лучей, которые хлорофиллом не поглощаются.
Фикобилины - красные и синие пигменты, содержащиеся у цианобактерий и некоторых водорослей. Исследования показали, что красные водоросли и цианобактерий наряду с хлорофиллом а содержат фикобилины. В основе химического строения фикобилинов лежат четыре пиррольные группировки.
Фикобилины представлены пигментами: фикоцианином, фикоэритрином и аллофикоцианином. Фикоэритрин - это окисленный фикоцианин. Фикобилины образуют прочные соединения с белками (фикобилинпротеиды). Связь между фикобилинами и белками разрушается только кислотой.
Фикобилины поглощают лучи в зеленой и желтой частях солнечного спектра. Это та часть спектра, которая находится между двумя основными линиями поглощения хлорофилла. Фикоэритрин поглощает лучи с длиной волны 495- 565 нм, а фикоцианин - 550- 615 нм. Сравнение спектров поглощения фикобилинов со спектральным составом света, в котором проходит фотосинтез у цианобактерий и красных водорослей, показывает, что они очень близки. Это позволяет считать, что фикобилины поглощают энергию света и, подобно каротиноидам, передают ее на молекулу хлорофилла, после чего она используется в процессе фотосинтеза. Наличие фикобилинов у водорослей является примером приспособления организмов в процессе эволюции к использованию участков солнечного спектра, которые проникают сквозь толщу морской воды (хроматическая адаптация). Как известно, красные лучи, соответствующие основной линии поглощения хлорофилла, поглощаются, проходя через толщу воды. Наиболее глубоко проникают зеленые лучи, которые поглощаются не хлорофиллом, а фикобилинами.
Свойства хлорофилла
Все хлорофиллы являются магниевыми солями пиррола. В центре молекулы хлорофилла находятся магний и четыре пиррольных кольца, соединенные друг с другом метановыми мостиками.
По химическому строению хлорофиллы - сложные эфиры дикарбоновой органической кислоты - хлорофиллина и двух остатков спиртов - фитола и метилового.
Важнейшей частью молекулы хлорофилла является центральное ядро. Оно состоит из четырех пиррольных пятичленных колец, соединенных между собой углеродными мостиками и образующих большое порфириновое ядро с атомами азота посередине, связанными с атомом магния. В молекуле хлорофилла есть дополнительное циклопентаноновое кольцо, которое содержит карбонильную, а также карбоксильную группы, связанные эфирной связью с метиловым спиртом. Наличие в порфириновом ядре конъюгированной по кругу системы десяти двойных связей и магния обусловливает характерный для хлорофилла зеленый цвет.
Хлорофилл в отличается от хлорофилла а только тем, что вместо метальной группы во втором пиррольном кольце имеет альдегидную группу СОН. Хлорофилла имеет сине-зеленую окраску, а хлорофилл в -- светло-зеленую. Адсорбируются они в разных слоях хроматограммы, что свидетельствует о разных химических и физических свойствах. По современным представлениям, биосинтез хлорофилла в идет через хлорофилл а.
Флуоресценция - это свойство многих тел под влиянием падающего света, в свою очередь, излучать свет: при этом длина волны излучаемого света обычно больше длины - волны возбуждающего света. Одним из важнейших свойств хлорофиллов является их ярко выраженная способность к флуоресценции, которая интенсивна в растворе и угнетена в хлорофилле, содержащемся в тканях листьев, в пластидах. Если смотреть на раствор хлорофилла в лучах света, проходящего через него, то он кажется изумрудно-зеленым, если же рассматривать его в лучах отраженного света, то он приобретает красную окраску - это явление флуоресценции.
Хлорофиллы различаются по спектрам поглощения, при этом у хлорофилла b по сравнению с хлорофиллом а полоса поглощения в красной области спектра несколько смещена в сторону коротковолновых лучей, а в сине-фиолетовой области максимум поглощения смещен в сторону длинноволновых (красных) лучей.
Фотосинтез
, образование зелеными растениями и некоторыми бактериями органических веществ с использованием энергии солнечного света. Происходит при участии (у растений ). В основе фотосинтеза лежат , в которых переносятся от донора (например, H 2 O, H 2 S) к акцептору (CO 2) с образованием восстановленных соединений (углеводов) и выделением O 2 (если донор электронов H 2 O), S (если донор электронов, например, H 2 S) и др.
Фотосинтез - один из самых распространенных процессов на Земле, обусловливает круговорот в природе углерода, O 2 и других элементов. Он составляет материальную и энергетическую основу всего живого на планете. Ежегодно в результате фотосинтеза в виде органических вещества связывается около 8·10 10 т углерода, образуется до 10 11 т . Благодаря фотосинтезу растения суши образуют около 1,8·10 11 т сухой биомассы в год; примерно такое же количество биомассы растений образуется ежегодно в Мировом океане. Тропический лес вносит до 29% в общую продукцию фотосинтеза суши, а вклад лесов всех типов составляет 68%. Фотосинтез высших растений и водорослей - единственный источник атмосферного O 2 .
Возникновение на Земле около 2,8 млрд. лет назад механизма с образованием O 2 представляет собой важнейшее событие в биологической эволюции, сделавшее свет Солнца главным источником свободной энергии , а воду - практически неограниченным источником водорода для синтеза веществ в живых организмах. В результате образовалась современного состава, O 2 стал доступным для пищи (см. ), а это обусловило возникновение высокоорганизованных гетеротрофных организмов (применяют в качестве источника экзогенные органические вещества).
Около 7% органических продуктов фотосинтеза человек использует в пищу, в качестве корма для животных, а также в виде и строительного материала. Ископаемое топливо - тоже продукт фотосинтеза. Его потребление в конце 20 в. примерно равно приросту биомассы.
Общее запасание энергии солнечного излучения в виде продуктов фотосинтеза составляет около 1,6·10 21 кДж в год, что примерно в 10 раз превышает современное энергетическое потребление человечества. Примерно половина энергии солнечного излучения приходится на видимую область спектра (длина волны l от 400 до 700 нм), которая используется для фотосинтеза (физиологически активная радиация, или ФАР). ИК излучение не пригодно для фотосинтеза кислородвыделяющих организмов (высших растений и водорослей), но используется некоторыми фотосинтезирующими бактериями.
Хлоропласт, представляющий собой замкнутую структуру, отделенную от остальной части клетки оболочкой, заключает в себе весь фотосинтетический аппарат. Световая стадия реализуется в мембранных структурах хлоропласта (так называемых тилакоидах), тогда как темновая стадия происходит в жидком содержимом хлоропласта (строме) при участии водорастворимых ферментов. У фотосинтезирующих бактерий хлоропласты отсутствуют, но световая стадия также осуществляется в мембранных образованиях - в так называемых хроматофорах.
Световая стадия фотосинтеза
Минимальная функциональная единица, еще способная осуществлять световую стадию фотосинтеза,- тилакоид. Он представляет собой микроскопический плоский диск, образованный белковолипидными мембранами, в которых находятся пигменты. В эти мембраны встроены все компоненты, необходимые для воды, никотинамиддинуклеотидфосфата (НАДФ) до НАДФН и синтеза АТФ из аденозиндифосфата. Световая стадия фотосинтеза инициируется поглощением кванта света пигментами, организованными в специальные светособирающие комплексы. Среди преобладает хлорофилл а
. К вспомогательным пигментам относятся хлорофилл b
, каротиноиды и др. Наличие светособирающей структуры из нескольких сотен или десятков молекул на каждый фотохимически активный (реакционный) центр на 2-3 порядка увеличивает сечение захвата излучения и обеспечивает возможность фотосинтеза при слабом освещении.
Часть вспомогательных пигментов, спектрально наиболее близких к фотохимически активному хлорофиллу, непосредственно окружает каждый из реакционных центров, образуя так называемые антенны.
Высокая эффективность переноса возбуждения от молекулы, поглотившей квант, к фотохимическому центру определяется спектральными свойствами и структурной организацией светособирающего комплекса и антенны, окружающей фотохимический центр. Эти пигменты обеспечивают передачу возбуждения за время менее 100 пс в пределах времени жизни синглетно возбужденного состояния хлорофилла.
В реакционном центре фотосинтеза, куда почти со 100%-ной вероятностью переносится возбуждение, происходит первичная реакция между фотохимически активной молекулой а
(у бактерий - бактериохлорофилла) и первичным акцептором электрона (ПА). Дальнейшие реакции в тилакоидных мембранах происходят между молекулами в их основных состояниях и не требуют возбуждения светом. Эти реакции организованы в электронтранспортную цепь - последовательность фиксированных в мембране переносчиков электрона. В электронтранспортной цепи высших растений и водорослей содержится два фотохимических центра (фотосистемы), действующих последовательно (рис. 2), в бактериальной электронтранспортной цепи - один (рис. 3).
В фотосистеме II высших растений и водорослей синглетно возбужденный хлорофилл а
в центре Р680 (число 680 обозначает, что максимум спектральных изменений системы при возбуждении светом находится вблизи 680 нм) отдает электрон через промежуточный акцептор к феофитину (ФЕО, безмагниевый аналог хлорофилла), образуя катион-радикал . Анион-радикал восстановленного феофитина служит далее донором электрона для связанного пластохинона (ПХ*; отличается от заместителями в хиноидном кольце), координированного с ионом Fe 3+ (в бактериях имеется аналогичный Fе 3+ -убихинонный комплекс). Далее электрон переносится по цепи, включающей свободный пластохинон (ПХ), присутствующий в избытке по отношению к остальным компонентам цепи, затем цитохромы (Ц) b 6
и f
, образующие комплекс с железо-серным центром, через медьсодержащий белок пластоцианин (ПЦ; мол. м. 10400) к реакционному центру фотосистемы I.
Центры быстро восстанавливаются, принимая электрон через ряд промежуточных переносчиков от воды. Образование O 2 требует последовательного четырехкратного возбуждения реакционного центра фотосистемы П и катализируется мембранным комплексом, содержащим Mn.
Хлорофилл a в фотосистеме I, имеющий максимум поглощения вблизи 700 нм (центр Р700), является первичным фотовозбуждаемым донором электрона, который он отдает первичному акцептору (ПА; его природа однозначно не установлена), а затем, через ряд промежуточных переносчиков (A i
) - растворимому белку ферредоксину (ФД), восстанавливающему с помощью фермента ферредоксин-НАДФ-редуктазы (ФНР) НАДФ до НАДФН. Катион-радикал окисленного пигмента восстанавливается пластоцианином.
В зрелых хлоропластах имеются граны (стопки тилакоидов), в мембранах которых присутствуют все компоненты злектронтранспортной цепи, и так называемые агранальные тилакоиды, не содержащие фотосистемы II.
Благодаря асимметрическому расположению компонентов электронтранспортной цепи относительно плоскости мембраны при разделении зарядов между хлорофиллом в каждом из двух фотосинтетических центров и акцептором электрона на тилакоидной мембране создается разность электрических потенциалов (плюс - на внутренней, минус - на внешней ее стороне). Перенос электрона пластохиноном сопровождается транспортом протонов, которые захватываются снаружи тилакоида при восстановлении пластохинона и освобождаются внутрь тилакоида при окислении пластогидрохинона. Перенос электронов сопряжен с синтезом АТФ из аденозиндифосфата (АДФ) и неорганического фосфата. Предполагают, что обратный транспорт протонов из тилакоидов в строму через белковый сопрягающий фактор (Н + -АТФ-синтетазу) сопровождается образованием .
Фотосистема I может действовать автономно без контакта с системой II. В этом случае циклический перенос электрона (на схеме показан пунктиром) сопровождается синтезом АТФ, а не НАДФН. Образующиеся в световой стадии коферменты НАДФН и АТФ используются в темновой стадии фотосинтез, в ходе которой снова образуется НАДФ и АДФ.
Электронтранспортные цепи фотосинтезирующих бактерий в основных своих чертах аналогичны отдельным фрагментам таковых в хлорогпастах высших растений. На рис. 3 показана электронтранспортная цепь пурпурных бактерий.
Темновая стадия фотосинтеза
Все фотосинтезирующие организмы, выделяющие O 2 , а также некоторые фотосинтезирующие бактерии сначала восстанавливают CO 2 до фосфатов Сахаров в так называемом Калвина. У фотосинтезирующих бактерий встречаются, по-видимому, и другие механизмы. Большинство цикла Калвина находится в растворимом состоянии в строме хлоропластов.
Упрощенная схема цикла показана на рис. 4. Первая стадия - карбоксилирование рибулозо-1,5-дифосфата и гидролиз продукта с ооразованием двух молекул 3-фосфоглицериновой кислоты. Эта С 3 -кислота фосфорилируется АТФ с образованием 3-фосфоглицероилфосфата, который затем восстанавливается НАДФН до глицеральдегид-3-фосфата. Полученный триозофосфат затем вступает в ряд реакций изомеризации, конденсации и перегруппировок, дающих 3 молекулы рибулозо-5-фосфата. Последний фосфорилируется при участии АТФ с образованием рибулозо-1,5-дифосфата и, таким образом, цикл замыкается. Одна из 6 образующихся молекул глицеральдегид-3-фосфата превращается в глюкозо-6-фосфат и используется затем для синтеза крахмала либо выделяется из хлоропласта в цитоплазму. Глицеральдегид-3-фосфат может также превращаться в 3-глицерофосфат и затем в липиды. Триозофосфаты, поступающие из хлоропласта, превращаются в основном в сахарозу, которая переносится из листа в другие части растения.
В одном полном обороте цикла Калвина расходуется 9 молекул АТФ и 6 молекул НАДФН для образования одной молекулы 3-фосфоглицериновой кислоты. Энергетическая эффективность цикла (отношение энергии фотонов, необходимых для фотосинтеза АТФ и НАДФН, к ΔG 0 образования углевода из CO 2) с учетом действующих в строме хлоропласта концентраций субстратов составляет 83%. В самом цикле Калвина нет фотохимических стадий, но световые стадии могут косвенно влиять на него (в том числе и на реакции, не требующие АТФ или НАДФН) через изменения концентраций ионов Mg 2+ и H + , а также уровня восстановленности ферредоксина.
Некоторые высшие растения, приспособившиеся к высокой интенсивности света и к теплому климату (например, сахарный тростник, кукуруза), способны предварительно фиксировать CO 2 в дополнительном С 4 -цикле. При этом CO 2 сначала включается в обмен четырехуглеродных дикарбоновых кислот, которые затем декарбоксилируются там, где локализован цикл Калвина. С 4 -цикл характерен для растений с особым анатомическим строением листа и разделением функций между двумя типами клеток: мезофильных, где сосредоточено карбоксилирование фосфоенолпировиноградной кислоты, и клеток обкладки сосудистого пучка, где функционирует цикл Калвина. Образующаяся в С 4 -цикле щавелевоуксусная кислота восстанавливается НАДФН до яблочной, которая перемещается в клетки сосудистой обкладки и здесь подвергается окислительному декарбоксилированию, образуя пировиноградную кислоту, CO 2 и НАДФН. Два последних используются в цикле Калвина, а пировиноградная кислота возвращается в С 4 -цикл (рис. 5). Физиологический смысл С 4 -цикла состоит в запасании CO 2 и повышении, таким образом, общей эффективности процесса.
Для кактусов, молочая и других засухоустойчивых растений характерно частичное разделение фиксации CO 2 и фотосинтеза во времени (CAM
-обмен, или обмен по типу толстянковых; CAM
сокр. от англ. Crassulaceae acid metabolism
). Днем устьица (каналы, через которые осуществляется газообмен с атмосферой) закрываются, чтобы уменьшить испарение воды. При этом поступление CO 2 также затруднено. Ночью устьица открываются, происходит фиксация CO 2 в виде фосфоенол-пировиноградной кислоты с образованием С 4 -кислот, которые днем декарбоксилируются, а освобождаемый при этом CO 2 включается в цикл Калвина (рис. 6).
Фотосинтез галобактерий
Единственный известный в природе нехлорофилльный способ запасания энергии света осуществляют бактерии Halobacterium halobium
. Ha ярком свету при пониженной O 2 они образуют в своих мембранах пурпурный белок . В результате индуцированной светом цис-транс
-изомеризации ретиналя (хромофора этого пигмента) происходит поглощение H + и синтез АТФ. Последний используется для частичного обеспечения энергетических потребностей клетки.
Фотосинтез: историческая справка
Ок. 1770 Дж.Пристли обнаружил, что растения выделяют O 2 . В 1779 Я.Ингенхауз установил, что для этого необходим свет и что O 2 выделяют только зеленые части растений. Ж.Сенебье в 1782 показал, что для питания растений требуется CO 2 ; в начале 19 в. H.Соссюр, исходя из закона сохранения массы, подтвердил, что большая часть массы растений создается из CO 2 и воды. В 1817 П.Пельтье и Ж.Каванту выделили зеленый пигмент хлорофилл. Позже К.А.Тимирязев показал близость спектра действия фотосинтеза и спектра поглощения хлорофилла. Ю.Сакс в середине 19 в., по-видимому, первым осознал, что этот продукт накапливается в хлоропластах, а Т.В.Энгельман доказал, что именно там же выделяется и O 2 .
В работах Ф.Блэкмана (1905), P.Эмерсона и У.Арнолда (1932), а также P.Хилла (1936-41) показано наличие световой и темновой стадий фотосинтеза и экспериментально реализована световая стадия в отсутствие CO 2 с использованием искусственных акцепторов электрона. Тем самым были получены подтверждения представлений об образовании O 2 путем воды. Окончательно это было доказано масс-спектрометрическим методом (С.Рубен, M.Камен, а также А.П.Виноградов и Р.В.Тейс, 1941).
В 1935-41 К.Ван Ниль обобщил данные по фотосинтезу высших растений и бактерий и предложил общее уравнение, охватывающее все типы фотосинтеза X.Гаффрон и К.Воль, а также Л.Дёйсенс в 1936-52 на основе количественных измерений выхода продуктов фотосинтеза поглощенного света и содержания сформулировали представление о "фотосинтетической единице" - ансамбле молекул пигмента, осуществляющих светосбор и обслуживающих фотохимический центр.
В 40-50-х гг. M.Калвин, используя изотоп 14 C, выявил механизм фиксации CO 2 . Д.Арнон (1954) открыл фотофосфорилирование (инициируемый светом синтез АТФ из АДФ и H 3 PO 4) и сформулировал концепцию электронного транспорта в мембранах хлоропластов. P.Эмерсон и Ч.M.Льюис (1942-43) обнаружили резкое снижение эффективности фотосинтеза при 700 нм (красное падение, или первый эффект Эмерсона), а в 1957 Эмерсон наблюдал неаддитивное усиление фотосинтеза при добавлении света низкой интенсивности с 650 нм к дальнему красному свету (эффект усиления, или второй эффект Эмерсона). На этом основании в 60-х гг. сформулировано представление о последовательно действующих фотосистемах в электронтранспортной цепи фотосинтеза с максимумами в спектрах действия вблизи 680 и 700 нм.
Основные закономерности образования O 2 при окислении в фотосинтезе установлены в работах Б.Кока и П.Жолио (1969-70). Близится к завершению выяснение молекулярной организации мембранного комплекса, катализирующего этот процесс. В 80-х гг. методом рентгеновского структурного анализа детально изучена структура отдельных компонентов фотосинтетического аппарата, включая реакционные центры и светособирающие комплексы (И.Дайзенхофер, X.Михель, P.Хубер).
Фотосинтез – это процесс трансформации поглощенной организмом энергии света в химическую энергию органических (и неорганических) соединений.
Процесс фотосинтеза выражают суммарным уравнением:
6СО 2 + 6Н 2 О ® С 6 Н 12 О 6 + 6О 2 .
На свету в зеленом растении из предельно окисленных веществ - диоксида углерода и воды образуются органические вещества, и высвобождается молекулярный кислород. В процессе фотосинтеза восстанавливаются не только СО 2 , но и нитраты или сульфаты, а энергия может быть направлена на различные эндэргонические процессы, в том числе на транспорт веществ.
Общее уравнение фотосинтеза может быть представлено в виде:
12 Н 2 О → 12 [Н 2 ] + 6 О 2 (световая реакция)
6 СО 2 + 12 [Н 2 ] → С 6 Н 12 О 6 + 6 Н 2 О (темновая реакция)
6 СО 2 + 12 Н 2 О → С 6 Н 12 О 6 + 6 Н 2 О + 6 О 2
или в расчете на 1 моль СО 2:
СО 2 + Н 2 О СН 2 О + О 2
Весь кислород, выделяемый при фотосинтезе, происходит из воды. Вода в правой части уравнения не подлежит сокращению, так как ее кислород происходит из СО 2 . Методами меченых атомов было получено, что Н 2 О в хлоропластах неоднородна и состоит из воды, поступающей из внешней среды и воды, образовавшейся в процессе фотосинтеза. В процессе фотосинтеза используются оба типа воды. Доказательством образования О 2 в процессе фотосинтеза служат работы голландского микробиолога Ван Ниля, который изучал бактериальный фотосинтез, и пришел к выводу, что первичная фотохимическая реакция фотосинтеза состоит в диссоциации Н 2 О, а не разложении СО 2 . Способные к фотосинтетической ассимиляции СО 2 бактерии (кроме цианобактерий) используют в качестве восстановителей Н 2 S, Н 2 , СН 3 и другие, и не выделяют О 2 . Такой тип фотосинтеза называется фоторедукцией:
СО 2 + Н 2 S → [СН 2 О] + Н 2 О + S 2 или
СО 2 + Н 2 А → [СН 2 О] + Н 2 О + 2А,
где Н 2 А – окисляет субстрат, донор водорода (у высших растений – это Н 2 О), а 2А – это О 2 . Тогда первичным фотохимическим актом в фотосинтезе растений должно быть разложение воды на окислитель [ОН] и восстановитель [Н]. [Н] восстанавливает СО 2 , а [ОН] участвует в реакциях освобождения О 2 и образования Н 2 О.
Солнечная энергия при участии зеленых растений и фотосинтезирующих бактерий преобразуется в свободную энергию органических соединений. Для осуществления этого уникального процесса в ходе эволюции был создан фотосинтетический аппарат, содержащий: I) набор фотоактивных пигментов, способных поглощать электромагнитное излучение определенных областей спектра и запасать эту энергию в виде энергии электронного возбуждения, и 2) специальный аппарат преобразования энергии электронного возбуждения в разные формы химической энергии. Прежде всего эторедокс-энергия, связанная с образованием высоковосстановленных соединений, энергия электрохимического потенциала, обусловленная образованием электрических и протонных градиентов на сопрягающей мембране (Δμ H +),энергия фосфатных связей АТФ и других макроэргических соединений, которая затем преобразуется в свободную энергию органических молекул.
Все эти виды химической энергии могут быть использованы в процессе жизнедеятельности для поглощения и трансмембранного переноса ионов и в большинстве реакций метаболизма, т.е. в конструктивном обмене.
Способность использовать солнечную энергию и вводить ее в биосферные процессы и определяет «космическую» роль зеленых растений, о которой писал великий русский физиологК.А. Тимирязев.
Процесс фотосинтеза представляет собой очень сложную систему по пространственной и временной организации. Использование высокоскоростных методов импульсного анализа позволили установить, что процесс фотосинтеза включает различные по скорости реакции - от 10 -15 с (в фемтосекундном интервале времени протекают процессы поглощения и миграции энергии) до 10 4 с (образование продуктов фотосинтеза). Фотосинтетический аппарат включает структуры с размерами от 10 -27 м 3 на низшем молекулярном уровне до 10 5 м 3 на уровне посевов.
Принципиальная схема фотосинтеза. Весь сложный комплекс реакций, составляющих процесс фотосинтеза, может быть представлен принципиальной схемой, в которой отображены основные стадии фотосинтеза и их сущность. В современной схеме фотосинтеза можно выделить четыре стадии, которые различаются по природе и скорости реакций, а также по значению и сущности процессов, происходящих на каждой стадии.
Физиология растений
Формула фотосинтеза
Две стадии фотосинтеза Процесс фотосинтеза состоит из двух последовательных и взаимосвязанных этапов: светового (фотохимического) и темнового (метаболического). На первой стадии происходит преобразование поглощенной фотосинтетическими пигментами энергии квантов света в энергию химических связей высокоэнергетического соединения АТФ и универсального восстановителя НАДФН - собственно первичных продуктов фотосинтеза, или так называемой «ассимиляционной силы». В темновых реакциях фотосинтеза происходит использование образовавшихся на свету АТФ и НАДФН в цикле фиксации углекислоты и ее последующего восстановления до углеводов. У всех фотосинтезирующих организмов фотохимические процессы световой стадии фотосинтеза происходят в особых энергопреобразующих мембранах, называемых тилакоидными, и организованы в так называемую электрон-транспортную цепь. Темновые реакции фотосинтеза осуществляются вне тилакоидных мембран (в цитоплазме у прокариот и в строме хлоропласта у растений). Таким образом, световая и темновая стадии фотосинтеза разделены в пространстве и во времени.
НАДФН – Никотинамидадениндинуклеотидфосфа́т
Синтез АТФ и образование НАДФН Одновременно с фотосинтетическим транспортом электронов происходит перенос протонов из стромы хлоропласта во внутритилакоидное пространство - возникает трансмембранный электрохимический градиент ионов водорода (pH-градиент), используемый затем комплексом фермента АТФ-синтазы для синтеза АТФ из АДФ и неорганического фосфата в процессе фотосинтетического фосфорилирования. При нециклическом токе электронов и сопряженном с ним фотофосфорилировании происходит образование восстановителя НАДФН и АТФ. При альтернативных путях переноса электронов - циклическом и псевдоциклическом - образуется только АТФ. Образующиеся в результате световых реакций первичные продукты фотосинтеза - НАДФН и АТФ - используются в ходе дальнейших ферментативных реакций для восстановления углекислоты до углеводов, жиров, белков. При неуглеводной направленности темнового метаболизма, когда преимущественно образуются аминокислоты, белки, органические кислоты, возрастает уровень потребления восстановителя НАДФН.
Темновые реакции фотосинтеза (фотосинтетическая фиксация CO 2) Метаболические варианты фотосинтетической фиксации CO 2 у растений принято классифицировать на С 3 -, С 4 - и САМ-фотосинтез. Образующиеся в темновых реакциях углеводы могут откладываться в виде крахмала в хлоропластах; выходить из хлоропластов и использоваться в образовании нового структурного материала клеток; служить источником энергии для различных метаболических процессов; транспортироваться в запасающие органы растения.
С 3 -путь фотосинтеза Восстановительный пентозофосфатный цикл фиксации CO 2 (С 3 -путь, или цикл Кальвина), открытый американскими учеными Э. Бенсоном и М. Калвином в 1950-е годы, универсален и обнаруживается практически у всех автотрофных организмов. В этом цикле (рис.5) фиксация СО 2 осуществляется на пятиуглеродное соединение рибулезобисфосфат (РуБФ) при участии фермента рибулезобисфосфаткарбоксилазы (РуБФ-карбоксилазы). Первым стабильным продуктом являются две молекулы трехуглеродного соединения 3-фосфоглицериновой кислоты (3-ФГК), восстанавливаемая затем с использованием АТФ и НАДФН до трехуглеводных сахаров, из которых образуется конечный продукт фотосинтеза - шестиуглеродная глюкоза. Субстратом ключевого фермента фотосинтетической фиксации СО 2 - РуБФ-карбоксилазы - наряду с СО 2 может быть и О 2 . При взаимодействии РуБФ с кислородом реализуется гликолатный, или С 2 -путь, известный как фотодыхание. Большинство наземных растений осуществляют фотосинтез по С 3 -пути. Типичные представители этой группы - горох, фасоль, конские бобы, шпинат, салат, капуста, пшеница, овес, рожь, ячмень, свекла, подсолнечник, тыква, томаты и другие одно- и двудольные растения.С 4 -путь фотосинтеза У некоторых видов растений (в основном тропических и очень небольшого числа видов из умеренных широт) первыми стабильными соединениями при фиксации СО 2 являются четырехуглеродные органические кислоты - яблочная и аспарагиновая. Такие растения отличаются видимым отсутствием фотодыхания (или очень низким уровнем), высокой скоростью фиксации СО 2 в расчете на единицу поверхности листа, более высокой общей фотосинтетической продуктивностью, быстрой скоростью роста. Функционально и анатомически в ткани их листьев выделяют 2 типа фотосинтезирующих клеток - клетки паренхимной обкладки, окружающие проводящие пучки, и клетки мезофилла. Для всех растений этой группы характерна катализируемая ферментом фосфоенолпируваткарбоксилазой (ФЕП-карбоксилазой) фиксация СО 2 на трехуглеродное соединение фосфоенолпируват (ФЕП) с образованием щавелевоуксусной кислоты, которая далее превращается в яблочную (малат) или аспарагиновую кислоту. Эти реакции протекают в цитоплазме клеток мезофилла листа. С 4 -кислоты затем поступают в клетки обкладки проводящих пучков, где подвергаются декарбоксилированию, а высвободившаяся СО 2 фиксируется через цикл Кальвина. Следовательно, у С 4 -растений фотосинтетический метаболизм углерода пространственно разделен и осуществляется в клетках различного типа, т. е. по «кооперативному механизму», подробно описанному австралийскими исследователями М. Хетчем и К Слэком и советским биохимиком Ю. С. Карпиловым в конце 1960-1970 годов. В соответствии с первичным механизмом декарбоксилирования С 4 -кислот все С 4 -растения подразделяются на три группы. НАДФ-малатдегидрогеназные растения осуществляют декарбоксилирование малата с помощью фермента НАДФ-малатдегидрогеназы в хлоропластах клеток обкладки проводящих пучков. Типичные представители этой группы - кукуруза, сахарный тростник, сорго, росичка кроваво-красная и другие злаки. НАД-малатдегидрогеназные растения осуществляют декарбоксилирование малата с помощью митохондриальной НАД-малатдегидрогеназы. Первичным продуктом фиксации углекислоты у них является аспартат. К типичным представителям этой группы принадлежат различные виды амаранта, портулак огородный, просо обыкновенное, бизонья трава, растущая в прериях Северной Америки и др. Фосфоенолпируват-карбоксикиназные растения осуществляют декарбоксилирование аспартата в цитоплазме клеток обкладки проводящих пучков с образованием ФЕП. Типичные представители - некоторые виды проса, хлориса, бутелуа. У суккулентных растений, произрастающих в условиях водного дефицита, фиксация СО 2 осуществляется с помощью так называемого САМ-пути (метаболизм кислот по типу растений семейства толстянковых). Первичный продукт фиксации углекислоты (яблочная кислота) образуется у них в темновой период и накапливается в вакуолях клеток листа. В дневное время при закрытых устьицах (которые закрываются для сохранения воды в тканях листа) осуществляется декарбоксилирование этой кислоты, а освобождающаяся СО 2 поступает в цикл Кальвина. Возникновение С4- и САМ-путей фотоассимиляции СО 2 связано с давлением на высшие наземные растения засушливого климата. С 4 -растения хорошо адаптированы к высокой интенсивности света, повышенным температурам и засухе. Оптимальная температура для осуществления фотосинтеза у них выше, чем у С 3 -растений. С 4 -растения наиболее многочисленны в зонах с высокими температурами. Они более экономно используют воду по сравнению с С 3 -растениями. В настоящее время известно, что все растения с С 4 -фотосинтезом - цветковые (из 19 семейств:16 - двудольных и 3 -однодольных). Не обнаружено ни одного семейства, которое бы состояло только из С 4 -растений.
Дыхание растений
Представляет процесс, соответствующий дыханию животных. Растение поглощает атмосферный кислород, а последний воздействует на органические соединения их тела таким образом, что в результате появляются вода и углекислота. Вода остается внутри растения, а углекислота выделяется в окружающую среду. При этом происходит уничтожение, трата органического вещества; следовательно, Д. прямо противоположно процессу ассимиляции углерода. До известной степени его можно уподобить окислению и горению вещества. Исходя из крахмала, схематическое уравнение Д. можно представить так:
C 6 H 10 O 5 (крахмал) +6O 2 (кислород) = 6CO 2 (углекислота) + 5H 2 O (вода)+ 2875 кДж/моль
Цикл Кребса. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
В анаэробных условиях пировиноградная кислота (пируват) подвергается дальнейшим превращениям в ходе спиртового, молочнокислого и других видов брожений, при этом НАДH используется для восстановления конечных продуктов брожения, регенерируя в окисленную форму. Последнее обстоятельство поддерживает процесс гликолиза, для которого необходим окисленный НАД + . В присутствии достаточного количества кислорода пируват полностью окисляется до С0 2 и Н 2 0 в дыхательном цикле, получившем название цикла Кребса , цикла ди- или трикарбоновых кислот . Все участки этого процесса локализованы в мАТФиксе или во внутренней мембране митохондрий.
Последовательность реакций в цикле Кребса. Участие органических кислот в дыхании давно привлекало внимание исследователей. Еще в 1910 г. шведский химик Т. Тунберг показал, что в животных тканях содержатся ферменты, способные отнимать водород от некоторых органических кислот (янтарной, яблочной, лимонной). В 1935 г. А. Сент-Дьердьи в Венгрии установил, что добавление к измельченной мышечной ткани небольших количеств янтарной, фумаровой, яблочной или щавелевоуксуснсй кислот резко активирует поглощение тканью кислорода.
Учитывая данные Тунберга и Сент-Дьердьи и исходя из собственных экспериментов по изучению взаимопревращения различных органических кислот и их влияния на дыхание летательной мышцы голубя, английский биохимик Г. А. Кребс в 1937 г. предложил схему последовательности окисления ди- и трикарбоновых кислот до С0 2 через «цикл лимонной кислоты» да счет отнятия водорода. Этот цикл и был назван его именем.
Непосредственно в цикле окисляется не сам пируват, а его производное -- ацетил-СоА. Таким образом, первым этапом на пути окислительного расщепления ПВК является процесс образования активного ацетила в ходе окислительного декарбоксилирования. Окислительное декарбоксилирование пирувата осуществляется при участии пируватдегидрогеназного мультиферментного комплекса. В состав его входят три фермента и пять коферментов. Коферментами служат тиаминпирофосфат (ТПФ) -- фосфорилированное производное витамина В ь липоевая кислота, коэнзим A, ФАД и НАД + . Пируват взаимодействует с ТПФ (декарбоксилазой), при этом отщепляется С0 2 и образуется гидроксиэтильное производное ТПФ (рис. 3). Последнее вступает в реакцию с окисленной формой липоевой кислоты. Дисульфидная связь липоевой кислоты разрывается и происходит окислительно-восстановительная реакция: гидроксиэтильная группа, присоединенная к одному атому серы, окисляется в ацетильную (при этом возникает высокоэнергетическая тиоэфирная связь), а другой атом серы липоевой кислоты восстанавливается. Образовавшаяся ацетиллипоевая кислота взаимодействует с коэнзимом А, возникают ацетил- СоА и восстановленная форма липоевой кислоты. Водород липоевой кислоты переносится затем на ФАД и далее на НАД + . В результате окислительного декарбоксилирования пирувата образуются ацетил-СоА, С0 2 и НАДH.
Рис. 3. Окислительное декарбоксилирование ПВК
Дальнейшее окисление ацетил-СоА осуществляется в ходе циклического процесса.
Цикл Кребса начинается с взаимодействия ацетил-СоА с енольной формой щавелевоуксусной кислоты. В этой реакции под действием фермента цитратсинтазы образуется лимонная кислота (2). Следующий этап цикла включает две реакции и катализируется ферментом аконитазой, или аконитатгидратазой (3). В первой реакции в результате дегидратации лимонной кислоты образуется цис- аконитовая. Во второй реакции аконитат гидратируется и синтезируется изолимонная кислота. Изолимонная кислота под действием НАД- или НАДФ-зависимой изоцитратдегидрогеназы (4) окисляется в нестойкое соединение -- щавелевоянтарную кислоту, которая тут же декарбоксилируется с образованием б-кетоглутаровой кислоты (б-оксоглутаровой кислоты).
б-Кетоглутарат, подобно пирувату, подвергается реакции окислительного декарбоксилирования. б-Кетоглутаратдегидрогеназный мультиэнзимный комплекс (5) сходен с рассмотренным выше пируватдегидрогеназным комплексом. В ходе реакции окислительного декарбоксилирования б-кетоглутарата выделяется С0 2 , образуются НАДH и сукцинил-СоА.
Подобно ацетил-СоА, сукцинил-СоА является высокоэнергетическим тиоэфиром. Однако если в случае с ацетил-СоА энергия тиоэфирной связи расходуется на синтез лимонной кислоты, энергия сукцинил-CoA может трансформироватся в образование фосфатной связи АТФ. При участии сукцинил- СоА-синтетазы (6) из сукцинил-СоА, АДФ и Н 3 Р0 4 образуются янтарная кислота (сукцинат), АТФ, регенерирует молекула СоА. АТФ образуется в результате субстратного фосфорилирования.
На следующем этапе янтарная кислота окисляется до фумаровой. Реакция катализируется сукцинатдегидрогеназой (7), коферментом которой является ФАД. Фумаровая кислота под действием фумаразы или фумаратгидратазы (8), присоединяя Н 2 0, превращается в яблочную кислоту (малат). И, наконец, на последнем этапе цикла яблочная кислота с помощью НАД- зависимой малатдегидрогеназы (9) окисляется в щавелевоуксусную. ЩУК, которая самопроизвольно переходит в енольную форму, реагирует с очередной молекулой ацетил-СоА и цикл повторяется снова.
Следует отметить, что большинство реакций цикла обратимы, однако ход цикла в целом практически необратим. Причина этого в том, что в цикле есть две сильно экзергонические реакции -- цитратсинтазная и сукцинил-СоА-синтетазная.
На протяжении одного оборота цикла при окислении пирувата происходит выделение трех молекул С0 2 , включение трех молекул Н 2 О и удаление пяти пар атомов водорода. Роль Н 2 О в цикле Кребса подтверждает правильность уравнения Палладина, который постулировал, что дыхание идет с участием Н 2 О, кислород которой включается в окисляемый субстрат, а водород с помощью «дыхательных пигментов» (по современным представлениям -- коферментов дегидрогеназ) переносится на кислород.
Энергетический выход цикла Кребса, его связь с азотным обменом. Цикл Кребса. играет чрезвычайно важную роль в обмене веществ растительного организма. Он служит конечным этапом окисления не только углеводов, но также белков, жиров и других соединений. В ходе реакций цикла освобождается основное количество энергии, содержащейся в окисляемом субстрате, причем большая часть этой энергии не теряется для организма, а утилизируется при образовании высокоэнергетических конечных фосфатных связей АТФ.
Каков же энергетический выход цикла Кребса? В ходе окисления пирувата имеют место 5 дегидрирований, при этом получаются 3НАДH, НАДФH (в случае изоцитратдегидрогеназы) и ФАДH 2 . Окисление каждой молекулы НАДH (НАДФH) при участии компонентов электронтранспортной цепи митохондрий дает по 3 молекулы АТФ, а окисление ФАДH 2 -- 2АТФ. Таким образом при полном окислении пирувата образуются 14 молекул АТФ. Кроме того, 1 молекула АТФ синтезируется; в цикле Кребса в ходе субстратного фосфорилирования. Следовательно, при окислении одной молекулы пирувата может образоваться 15 молекул АТФ. А поскольку в процессе гликолиза из молекулы глюкозы возникают две молекулы пирувата, их окисление даст 30 молекул АТФ.
Итак, при окислении глюкозы в процессе дыхания при функционировании гликолиза и цикла Кребса в общей сложности образуются 38 молекул АТФ (8 АТФ связаны с глико- лизом). Если принять, что энергия третьей сложноэфирнои фосфатной связи АТФ равняется 41,87 кДж/моль (10 ккал/моль), то энергетический выход гликолитического пути аэробного дыхания составляет 1591 кДж/моль (380 ккал/моль).
Значение цикла Кребса не ограничивается его вкладом в энергетический обмен клетки. Не менее важную роль играет то обстоятельство, что многие промежуточные продукты цикла используются при синтезе различных соединений. Из кетокислот в ходе реакций переаминирования образуются аминокислоты. Для синтеза липидов, полиизопренов, углеводов и ряда других соединений используется ацетил-СоА.
Регуляция цикла Кребса. Дальнейшее использование образующегося из пирувата ацетил-СоА зависит от энергетического состояния клетки. При малой энергетической потребности клетки дыхательным контролем тормозится работа дыхательной цепи, а следовательно, реакций ЦТК и образования интермедиатов цикла, в том числе оксалоацетата, вовлекающего ацетил-СоА в цикл Кребса. Это приводит к большему использованию ацетил-СоА в синтетических процессах, которые также потребляют энергию.
Особенностью регуляции ЦТК является зависимость всех четырех дегидрогеназ цикла (изоцитратдегидрогеназы, б-кетоглутаратдегидрогеназы, сукцинатдегидрогеназы, малатдегидрогеназы) от отношения [НАДH]/[НАД + ]. Активность цитратсинтазы тормозится высокой концентрацией АТФ и собственным продуктом -- цитратом. Изоцитратдегидрогеназа ингибируется НАДH и активируется цитратом. б-Кето- глутаратдегидрогеназа подавляется продуктом реакции -- сукцинил-СоА и активируется аденилатами. Окисление сукцината сукцинатдегидрогеназой тормозится оксалоацетатом и ускоряется АТФ, АДФ и восстановленным убихиноном (QH 2). Наконец, малатдегидрогеназа ингибируется оксалоацетатом и у ряда объектов -- высоким уровнем АТФ. Однако степень участия величины энергетического заряда, или уровня адениновых нуклеотидов, в регуляции активности цикла Кребса у растений до конца не выяснена.
Регулирующую роль может играть также альтернативный путь транспорта электронов в растительных митохондриях. В условиях высокого содержания АТФ, когда активность основной дыхательной цепи снижена, окисление субстратов через альтернативную оксидазу (без образования АТФ) продолжается, что поддерживает на низком уровне отношение НАДH/НАД + и снижает уровень АТФ. Все это позволяет циклу Кребса функционировать.
|
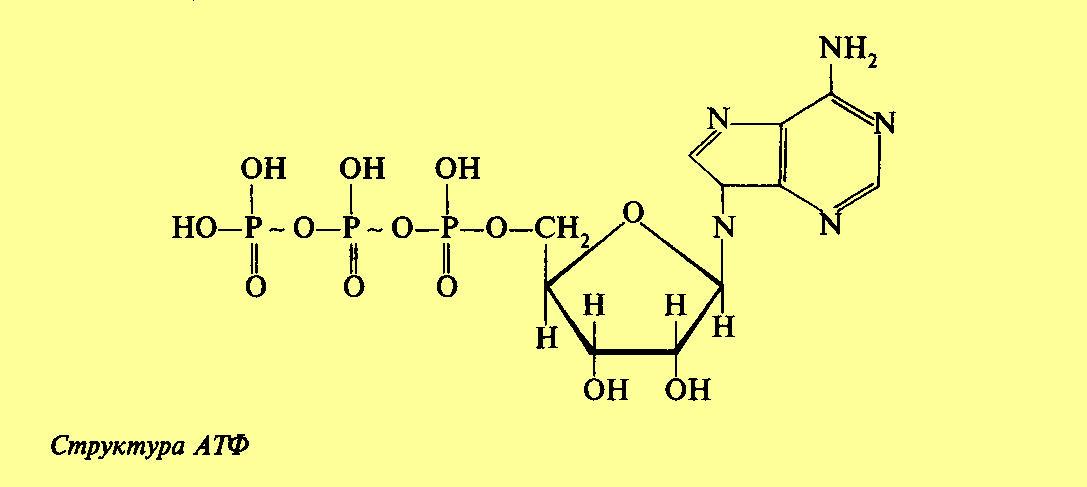
Аденозинтрифосфат. Структура и функции |
|
Процессы обмена вещества включают в себя реакции, идущие с потреблением энергии, и реакции с выделением энергии. В некоторых случаях эти реакции сопряжены. Однако часто реакции, в которых энергия выделяется, отделены в пространстве и во времени от реакций, в которых она потребляется. В процессе эволюции у растительных и животных организмов выработалась возможность хранения энергии в форме соединений, обладающих богатыми энергией связями. Среди них центральное место занимает аденозинтрифосфат (АТФ). АТФ представляет собой нуклеотидфосфат, состоящий из азотистого основания (аденина), пентозы (рибозы) и трех молекул фосфорной кислоты. Две концевые молекулы фосфорной кислоты образуют макроэргические, богатые энергией связи. В клетке АТФ содержится, главным образом, в виде комплекса с ионами магния. Аденозинтрифосфйт в процессе дыхания образуется из аденозиндифосфата и остатка неорганической фосфорной кислоты (Ф н) с использованием энергии, освобождающейся при окислении различных органических веществ: АДФ + Ф н -> АТФ + Н 2 0. При этом энергия окисления органических соединений превращается в энергию фосфорной связи. В 1939-1940 гг. Ф. Липман установил, что АТФ служит главным переносчиком энергии в клетке. Особые свойства этого вещества определяются тем, что конечная фосфатная группа легко переносится с АТФ на другие соединения или отщепляется с выделением энергии, которая может быть использована на физиологические функции. Эта энергия представляет собой разность между свободной энергией АТФ и свободной энергией образующихся продуктов (AG). AG - это изменение свободной энергии системы или количество избыточной энергии, которая освобождается при реорганизации химических связей. Распад АТФ происходит по уравнению: АТФ + Н 2 0 -> АДФ + Ф н, при этом
происходит как бы разрядка аккумулятора, при рН = 7 выделяется AG = -30,6 кДж. Этот процесс катализируется ферментом аденозинтрифосфатазой (АТФаза). Равновесие гидролиза АТФ смещено в сторону завершения реакции, что и обусловливает большую отрицательную величину свободной энергии гидролиза. Это связано с тем, что при диссоциации четырех гидроксильных группировок при рН = 7 АТФ имеет четыре отрицательных заряда. Близкое расположение зарядов друг к другу способствует их отталкиванию и, следовательно, отщеплению фосфатных группировок. В результате гидролиза образуются соединения с одноименным зарядом (АДФ 3- и НР0 4 2-), которые отталкиваются друг от друга, что препятствует их соединению. Уникальные свойства АТФ объясняются не только тем, что при ее гидролизе выделяется большое количество энергии, но и тем, что она обладает способностью отдавать концевую фосфатную группу вместе с запасом энергии на другие органические соединения. Энергия, заключенная в макроэргической фосфорной связи, используется на физиологическую деятельность клетки. Вместе с тем по величине свободной энергии гидролиза - 30,6 кДж/моль АТФ занимает промежуточное положение. Благодаря этому система АТФ - АДФ может служить переносчиком фосфатных групп от фосфорных соединений с более высокой энергией гидролиза, например фосфоенолпируват (53,6 кДж/моль), к соединениям с более низкой энергией гидролиза, например сахарофосфатам (13,8 кДж/моль). Таким образом, система АТФ - АДФ является как бы промежуточной или сопрягающей. |
|
С химической точки зрения дыхание - это медленное окисление. При окислительно-восстановительных реакциях происходит перенос водорода или электрона от донора ДН 2 (который окисляется) к акцептору А (который восстанавливается): ДН 2 +А ->Д + АН 2 . Для того чтобы судить о направлении движения электронов между какими-либо двумя веществами, вводится понятие стандартного восстановительного потенциала (Е 0) - это мера электронного давления. За нуль потенциала условно принят восстановительный потенциал реакции Н 2 -> 2Н + + 2е. Чем более отрицательна величина восстановительного потенциала, тем больше способность данного вещества отдавать электроны (окисляться) или служить восстановителем. Наоборот, чем положительнее величина восстановительного потенциала данного вещества, тем больше его способность воспринимать электроны (восстанавливаться или служить окислителем). Восстановительный потенциал кислорода равен +0,81В. В создании современных представлений о биологическом окислении большое значение имели работы двух крупнейших русских ученых - В.И. Палладина (1859-1922) и А.Н. Баха (1857-1946). Работы А.Н. Баха были посвящены возможности активации кислорода воздуха. Молекулярный кислород - достаточно инертное соединение. Бах выдвинул предположение, что имеются ферменты - оксигеназы, активирующие кислород. Он считал, что процесс активации состоит в том, что происходит образование пероксидных соединений. В.И. Палладии впервые стал рассматривать дыхание как ряд ферментативных реакций. Основное значение в процессе окисления он придавал процессу отнятия водорода от субстрата при участии воды. Содержание своей теории В.И. Палладии выразил в виде следующих уравнений: С 6 Н 12 0 6 + 6Н 2 0 + 12R -> 6С0 2 + 12RH 2 + 6O 2 -> 12R + 12Н 2 0 С 6 Н 12 0 6 + 60 2 - 4 6С0 2 + 6Н 2 0 Символом R В.И. Палладии обозначал дыхательный пигмент, способный к обратимым окислительно-восстановительным превращениям. Из приведенной схемы вытекают следующие важные положения: 1. Непременным участником дыхания является вода. 2. Вода наряду с окисляемым субстратом выполняет роль донора водорода. 3. В процессе дыхания участвуют специфические активаторы водорода, отнимающие водород от субстрата. 4. Первые этапы дыхания являются анаэробными и не требуют присутствия молекулярного кислорода. 5. Молекулярный кислород используется на заключительном этапе дыхания для регенерации акцепторов водорода с образованием воды. Все указанные положения легли, как мы увидим, в основу современных представлений о процессе дыхания, согласно которым дыхание происходит в две фазы - анаэробную и аэробную, и молекулярный кислород используется на регенерацию ферментов за счет Н+ воды и субстрата. В процессе дыхания активируется как водород субстрата, так и кислород воздуха. |
|
Пути дыхательного обмена |
|
Существуют две основные системы и два основных пути превращения дыхательного субстрата, или окисления углеводов: 1) гликолиз + цикл Кребса (гликолитический); 2) пентозофосфатный (апотомический). Относительная роль этих путей дыхания может меняться в зависимости от типа растений, возраста, фазы развития, а также в зависимости от факторов среды. Процесс дыхания растений осуществляется во всех внешних условиях, при которых возможна жизнь. Растительный организм не имеет приспособлений к регуляции температуры, поэтому процесс дыхания осуществляется при температуре от -50 до +50°С. Нет приспособлений у растений и к поддержанию равномерного распределения кисло¬рода по всем тканям. Именно необходимость осуществления процесса дыхания в разнообразных условиях привела к выработке в процессе эволюции разнообразных путей дыхательного обмена и к еще большему разнообразию ферментных систем, осуществляющих отдельные этапы дыхания. При этом важно отметить взаимосвязь всех процессов обмена в организме. Изменение пути дыхательного обмена приводит к глубоким изменениям во всем метаболизме растений. |
Данный путь дыхательного обмена является наиболее распространенным и, в свою очередь, состоит из двух фаз. Первая фаза - анаэробная (гликолиз), вторая фаза -аэробная . Эти фазы локализованы в различных компартментах клетки. Анаэробная фаза гликолиз - в цитоплазме, аэробная фаза - в митохондриях. Обычно химизм дыхания начинают рассматривать с глюкозы. Вместе с тем в растительных клетках глюкозы мало, поскольку конечными продуктами фотосинтеза являются сахароза как основная транспортная форма сахара в растении или запасные углеводы (крахмал и др.). Поэтому, чтобы стать субстратом дыхания сахароза и крахмал должны гидролизоваться с образованием глюкозы.
-
 17 апреля 2015Соляная лампа «Каменный цветок
17 апреля 2015Соляная лампа «Каменный цветок -
 17 апреля 2015Продуктивные способы потратить время в интернете
17 апреля 2015Продуктивные способы потратить время в интернете -
 17 апреля 2015Как установить приложение плей маркет на ноутбук
17 апреля 2015Как установить приложение плей маркет на ноутбук





 Скрытый бан аккаунта в instagram")
